El ojo es un órgano fascinante debido a la extremada perfección de su diseño. Mediante él, percibimos los cambios de color que ocurren en la naturaleza; nos adaptamos a ambientes con iluminación muy intensa (como ocurre en el día), o bien, casi imperceptible (como ocurre en la obscuridad o en la noche). Estas capacidades se deben a la presencia de las células fotoreceptoras en la retina, capaces de detectar paquetes individuales o cuantos de luz. De manera adicional, nosotros poseemos un par de lentes (Medios ópticos con alto poder de refracción): la córnea y el cristalino, que enfocan la luz captada por la cámara anterior del ojo, y llevan a la formación de imágenes en la retina.
Podemos decir que los rasgos distintivos del ojo de los vertebrados son: i) la íntima integración de los diferentes tejidos que lo constituyen; ii) el control de su desarrollo mediante un grupo de genes que se han conservado a lo largo de la evolución, desde invertebrados como las medusas hasta organismos tan complejos como los insectos o los mamíferos. iii) Finalmente, para que se forme un órgano tan especializado como el que encontramos en el adulto, se requiere una serie de interacciones tisulares complejas.
Por esto, el ojo es uno de los focos de atención en la Biología del Desarrollo. A principios del Siglo XX, Hans Spemann e Hilde Mangold realizaron experimentos de ablación y recombinación de tejidos que en conjunto, condujeron a una serie de conceptos clave en el desarrollo del ojo, y que sentaron las bases para que ellos propusieran el fenómeno de inducción embrionaria. De acuerdo con sus resultados y los de otros autores, la formación del cristalino depende de la interacción entre la vesícula óptica y el ectodermo cefálico. Del mismo modo, los Biólogos del Desarrollo han propuesto que este tipo de interacciones entre el ectodermo superficial y el cristalino son requeridas para la formación de la retina. A pesar de la existencia de datos contradictorios en estos experimentos, actualmente los científicos aceptan el hecho de que el desarrollo de estructuras complejas como el ojo depende de interacciones tisulares (ver Figura 1).
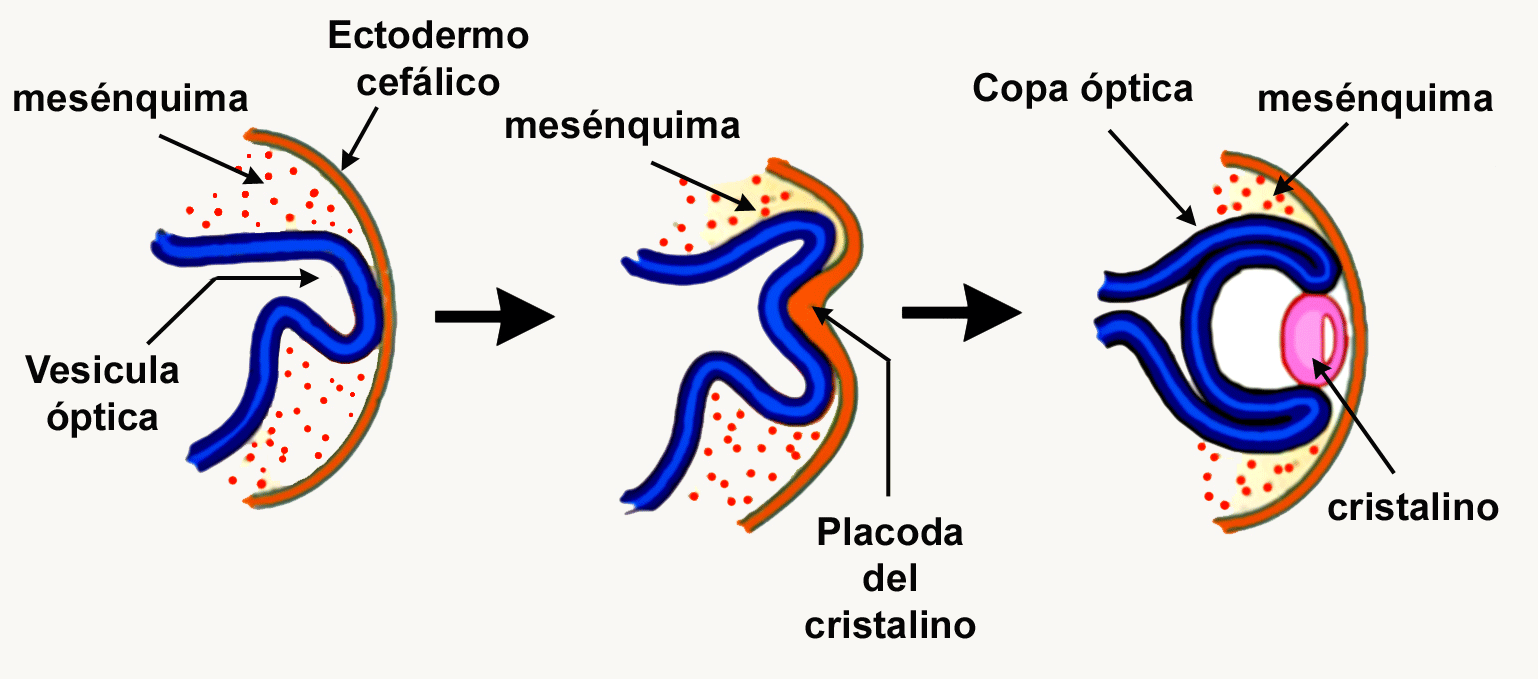
Figura 1. Esquema que ilustra las etapas iniciales del desarrollo del ojo de los mamíferos. Hasta ahora se pensaba que la formación de esta estructura dependía de interacciones inductivas en las que participaban diferentes tejidos embrionarios; principalmente el ectodermo cefálico, el neuroectodermo que evaginaba del tubo neural y en etapas posteriores el cristalino. Estas interacciones, aparentemente, determinaban la aparición de la retina funcional.
No obstante, en la revista Nature del 7 de abril de 2011 (1) un grupo de investigadores del centro RIKEN para Biología del Desarrollo de Kobe, Japón, e investigadores de las Universidades de Kyoto y de Osaka, también en Japón, publicaron un trabajo altamente sobresaliente. Este trabajo, se basa en el cultivo de células troncales de embrión de ratón en suspensión bajo condiciones en las que los investigadores no añadieron suero de origen animal al sistema experimental (SFEBq). Estas condiciones fueron descritas por el mismo grupo en 2008 (2); en esa ocasión, ellos señalaron que la expresión de fenotipos particulares, principalmente neuronas del tipo encontrado en neonatos, podía inducirse con aditivos particulares como son algunos componentes de matriz extracelular y algunos factores de crecimiento (2).
Bajo estas condiciones, el grupo de Yoshiki Sasai cultivó células troncales de embrión de ratón, e incluyó un tratamiento temporal con activina. Siguiendo este esquema experimental, ellos lograron replicar el proceso de diferenciación retinal. Más aún, estos investigadores añadieron al medio de cultivo una mezcla de componentes de matriz extracelular (Matrigel), o bien utilizaron laminina y entactina purificadas, y obtuvieron una estructura epitelial rígida y continua que al ser mantenida en cultivo por períodos prolongados se organizaba en una estructura similar a la copa óptica. El proceso se presentó en 4 etapas morfológicas muy similares a las observadas en el embrión de ratón (Figura 2). De manera adicional, ellos encontraron que esta estructura presentó dos capas bien definidas, y muy similares a las encontradas durante el desarrollo ocular: una porción distal, que acumula pigmento y corresponde al epitelio retinal pigmentario, y una porción proximal, cóncava, con varias capas de células neuronales y que correspondería al tejido fotorreceptor de la retina. Estas similitudes fueron tanto morfológicas como moleculares, y en parte, funcionales.
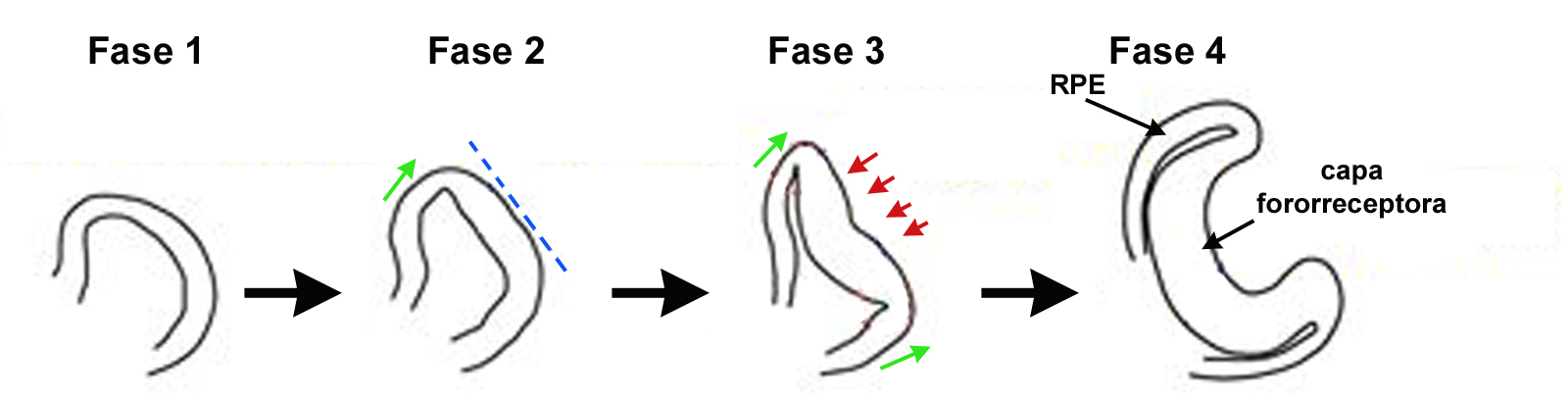
Figura 2. Fases del desarrollo de la estructura retinal obtenida a partir de agregados celulares similares a cuerpos embrioides, formados por células troncales de ratón cultivadas en medio sin suero, en presencia de activina y Matrigel, o laminina y entactina purificadas.En la fase 2, se inicia la migración lateral de la evaginación inicial, seguida popr su aplanamiento (línea punteada azul). En la fase 3 y 4 se establecen las dos capas de la copa óptica. La porción externa, distal, se diferencia en epitelio pigmentario retinal (RPE), y la capa interna, proxcimal, se constituye en el elemento fotorreceptor. Flechas rojas indican procesos de invaginación; verdes, evaginación.
De manera importante, con estos experimentos Sasai y sus colaboradores demostraron que el proceso de desarrollo de la retina se encuentra formando parte del programa de desarrollo, y al contrario de lo que se suponía, no parece depender de señales o interacciones inductivas. En sí, el proceso parece ser dirigido por la expresión del gene homeótico Rx (3). Por otra parte, mediante el uso de inhibidores de la actividad de miosina, los investigadores concluyeron que la formación de la retina depende en sus dos primeras fases de esta proteína, mientras que las fases 3 y 4 son mucho más dependientes de la actividad proliferativa de la población celular.
La metodología que Sasai y sus colaboradores desarrollaron, aunque permite obtener una estructura tridimensional y diferenciada, todavía no conduce a la obtención de una retina funcional ya que falta establecer las conexiónes fisiológicas con un sistema nervioso. Además, la retina generada in vitro no posee un número suficiente de elementos fotorreceptores (conos y bastones). No obstante, este modelo abre la posibilidad de generar después de varios años de investigación, sistemas in vitro para el modelaje experimental de enfermedades de la retina, y para el desarrollo y ensayo de drogas oftalmológicas. Quizás a muy largo plazo nos permita generar retinas sintéticas a partir de las cuales podamos aislar precursores de las células fotorreceptoras para emplearlos en el transplante terapéutico.
Lecturas sugeridas:1. Eiraku M, Takata N, Ishibashi H, Kawada M, Sakakura E, Okuda S, Sekiguchi K, Adachi T, Sasai Y. (2011).Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature 472:51-56.
2. Eiraku M, Watanabe K, Matsuo-Takasaki M, Kawada M, Yonemura S, Matsumura M, Wataya T, Nishiyama A, Muguruma K, Sasai Y. (2008) Self-organized formation of polarized cortical tissues from ESCs and its active manipulation by extrinsic signals. Cell Stem Cell. 3:519-532.
3. Mathers PH., Grinberg A., Mahon KA., Jamrich M. (1997). The Rx homeobox gene is essential for vertebrate eye development. Nature 387:603-607..
|